






Sán lá ruột nhỏ Haplorchis taichui (Nishigori, 1924) thuộc họ Heterophyidae, lớp Trematoda, ngành Platyhelminthes, kí sinh và gây bệnh ở người và động vật. Toàn bộ hệ gen ty thể (mitochondrial DNA genome, mtDNA) của H. taichui, chủng QT (HTAQT, Quảng Trị) đã được xác định cấu trúc và đặc điểm gen học, dữ liệu có giá trị phục vụ nghiên cứu dịch tễ học phân tử, thành phần loài, chẩn đoán, phân loại và phòng chống bệnh. Hệ gen mtDNA chủng HTAQT có độ dài 15.119 bp, chứa 36 gen, trong đó có 12 gen mã hóa protein (cox1, cox2, cox3, nad1, nad2, nad3, nad4L, nad4, nad5, nad6, atp6 và cob); 2 gen RNA ribosome (rRNA) gồm rrnL (16S) và rrnS (12S); 22 gen vận chuyển RNA (tRNA hay trn); và một vùng không mã hóa (non-coding, NR), chia thành 2 tiểu vùng là không mã hóa ngắn (SNR) và không mã hóa dài (LNR). Vùng LNR, độ dài 1.692 bp, nằm giữa vị trí của trnG (vận chuyển Glycine) và trnE (Glutamic acid), chứa 6 cấu trúc lặp liền kề kế tiếp nhau (tandem repeat, TR), sắp xếp lần lượt là: TR1A, TR2A, TR1B, TR2B, TR3A, TR3B. Từng gen mã hóa protein (12 gen), rRNA ribosome (2 gen) và tRNA vận chuyển (22 gen) đã được phân tích, cụ thể gen mã hóa protein được xác định kích thước, cách sử dụng bộ mã khởi đầu và kết thúc; các gen rRNA và tRNA được xác định cấu trúc bậc hai.
MỞ ĐẦU
Bệnh sán lá ruột nhỏ (haplorchiasis) ở người chủ yếu do loài sán lá ruột nhỏ Haplorchis taichui và H. pumilio gây nên, được phát hiện phổ biến ở các nước châu Á, một phần châu Phi và Trung Nam Mỹ (Chai et al., 2009; 2010; Dung et al., 2007). Tại Việt Nam, do tập quán ăn gỏi cá nên tỷ lệ người mắc bệnh kí sinh trùng truyền qua thực phẩm rất cao, trong đó có bệnh do Haplorchis spp gây ra. Sán lá ruột nhỏ H. taichui và H. pumilio thuộc giống Haplorchis trong họ Heterophyidae, H. taichui được Nishigori phát hiện lần đầu tiên vào năm 1924, ký sinh trên người ở Philippin. Cho đến nay, H. taichui, H. pumilio được ghi nhận là lưu hành rộng rãi ở nhiều quốc gia trên thế giới trong đó có Việt Nam (Dung et al., 2007; Chai et al., 2012; De, Le, 2011; Nguyễn Thị Khuê et al., 2015). Tại Việt Nam, haplorchiasis do H. taichui và H. pumilio được phát hiện ở hầu hết các tỉnh thành trong cả nước, đặc biệt ở các tỉnh phía Bắc và miền Trung (Dung et al., 2007; Kim Văn Vạn et al., 2007; Nguyễn Thị Khuê et al., 2015).
Hệ gen ty thể (mitochondrial genome) của sán dẹt (platyhelminth) có cấu trúc là một vòng DNA hai sợi khép kín, kích thước khoảng 13.5- – 21 kb, chứa một hệ gen riêng gồm 12 gen mã hóa cho sản phẩm protein, 2 gen RNA ribosome (rRNA), 22 gen RNA vận chuyển (tRNA hoặc trn) và một phần không mã hóa (non-coding region, NR) dài ngắn khác nhau (Le et al., 2002). Nghiên cứu giải mã hệ gen ty thể và phân tích đặc điểm của từng gen riêng biệt, đang được quan tâm và thực hiện trên toàn thế giới, nhằm mục đích để giải đáp các câu hỏi về chức năng, tiến hoá, nguồn gốc, phả hệ và di truyền quần thể (Bernt et al., 2013). Chính vì vậy, đã có hàng chục hệ gen ty thể của sán lá đã được giải mã thành công và được ứng dụng hiệu quả trong nghiên cứu bệnh động vật lây sang người (Le et al., 2002; Jia et al., 2012).
Nhằm cung cấp dữ liệu cấu trúc hệ gen ty thể và đặc điểm gen học các loài sán lá ruột nhỏ đang kí sinhXác định cấu trúc và đặc điểm gen học hệ gen ty thể của sán lá ruột nhỏ Haplorchis taichui (Trematoda: Heterophyidae) và gây bệnh trên người và động vật chúng tôi tiến hành giải trình tự và phân tích đặc điểm gen của một số kí sinh trùng thường gặp tại Việt Nam, thuộc các đề tài NAFOSTED (giai đoạn 2010- – 2016). Trong bài báo này, hệ gen ty thể của loài sán lá ruột nhỏ H. taichui, mẫu thu thập tại Quảng Trị, Việt Nam (kí hiệu HTAQT) đã được thu nhận và phân tích đầy đủ cung cấp dữ liệu cho các hướng nghiên cứu khác nhau sử dụng chuỗi gen/chỉ thị phân tử có từ hệ gen ty thể.
NGUYÊN LIỆU VÀ PHƯƠNG PHÁP
Mẫu Haplorchis taichui trong nghiên cứu và tách chiết DNA tổng số
Mẫu sán lá ruột nhỏ H. taichui trưởng thành thu từ một bệnh nhân tại Quảng Trị (Việt Nam), ký hiệu HTAQT, được sử dụng để giải trình tự hệ gen ty thể. Mẫu đã được xác định loài dựa vào đặc điểm hình thái học (De, Le, 2011) và khẳng định bằng sinh học phân tử sử dụng chỉ thị cox1 và phân tích phả hệ xác định quan hệ về loài (Nguyễn Thị Khuê et al., 2015).
DNA tổng số được tách chiết bằng bộ sinh phẩm bộ sinh phẩm DNeasy Blood and Tisue kit (hãng Qiagen) theo hướng dẫn của nhà sản xuất, như đã sử dụng trước đây (Nguyễn Thị Khuê et al., 2015). DNA tổng số của mẫu được bảo quản ở – 20oC và khuôn DNA tổng số được pha ở nồng độ 50 ng/μl, sử dụng cho phản ứng PCR (1-2 μl/phản ứng).
Thiết kế mồi và thực hiện PCR
Trước tiên, một số cặp mồi cơ bản được thiết kế dựa vào so sánh trình tự của các gen chọn lọc lấy từ các hệ gen ty thể của các loài sán dẹt có trong Ngân hàng gen (Le et al., 2002); để thực hiện PCR dài (long, LPCR) thu nhận đoạn DNA hệ gen ty thể kích thước 3-8 kb và PCR đơn thuần thu PCR sản phẩm dưới 3 kb.
Bảng 1 liệt kê tất cả mồi chung (14 mồi đầu tiên trong danh sách) và mồi đặc hiệu (17 mồi còn lại) được thiết kế và sử dụng. Sau đó trên cơ sở trình tự nucleotide của các chuỗi đã thu nhận tiếp tục thiết kế mồi và thực hiện giải trình tự bằng phương pháp “lao mồi” (primerwalking); hoặc/và sau khi tách dòng. PCR được thực hiện với chu trình nhiệt gồm các bước 94oC/5 phút trong 1 chu kỳ; tiếp theo là 35 chu kỳ ở 94oC/1 phút, 50oC/1 phút và 72oC/1-4 phút; chu kỳ cuối kéo dài 10 phút ở 72oC. Đối với LPCR, thực hiện bước kéo dài ở 72oC/6–10 phút. Sản phẩm PCR được tinh sạch bằng QIAquick PCR Purification Kit (QIAGEN) và được dòng hoá vào vector pCR2.1 hoặc pCR2.1-TOPO TAcloning Kit (Invitrogen), chuyển nạp vào dòng tế bào IVNαF’ để chọn lọc khuẩn lạc. DNA của plasmid tái tổ hợp được tách chiết với bộ hoá chất QIAprep Spin Plasmid Extraction Kit (QIAGEN).
Giải trình tự và phân tích chuỗi gen
Trình tự nucleotide từ giải trình tự trực tiếp sản phẩm PCR hoặc từ DNA plasmid tái tổ hợp trên máy tự động ABI Avant 3100 Genetic Analyzer (Mỹ), được xử lý bằng chương trình SeqEd1.3; sau đó so sánh sử dụng chương trình AssemblyLIGN1.9 và MacVector8.2 (Accelrys Inc.). Xác định cấu trúc gen bằng phần mềm MacVector8.2 và GENEDOC2.7 (http://iubio.bio.indiana.edu/soft/molbio/ibmpc/genedo c-readme.html), xác định cấu trúc RNA vận chuyển bằng chương trình tRNAscan-SE1.21 (http://lowelab.ucsc.edu/tRNAscan-SE/) hoặc và chương trình ARWEN (http://mbioserv2. mbioekol.lu.se/ARWEN/), cấu trúc của RNA ribosome sử dụng chương trình RNAviz (http://rnaviz.sourceforge.net/). Trình tự amino acid từ trình tự nucleotide các gen mã hóa protein của hệ gen ty thể được suy diễn sử dụng bộ mã của sán dẹt (flatworm mitochondrial code) có trong chương trình GENEDOC2.7.
KẾT QUẢ VÀ THẢO LUẬN
Cấu trúc và sắp xếp hệ gen ty thể của sán lá ruột nhỏ H. taichui
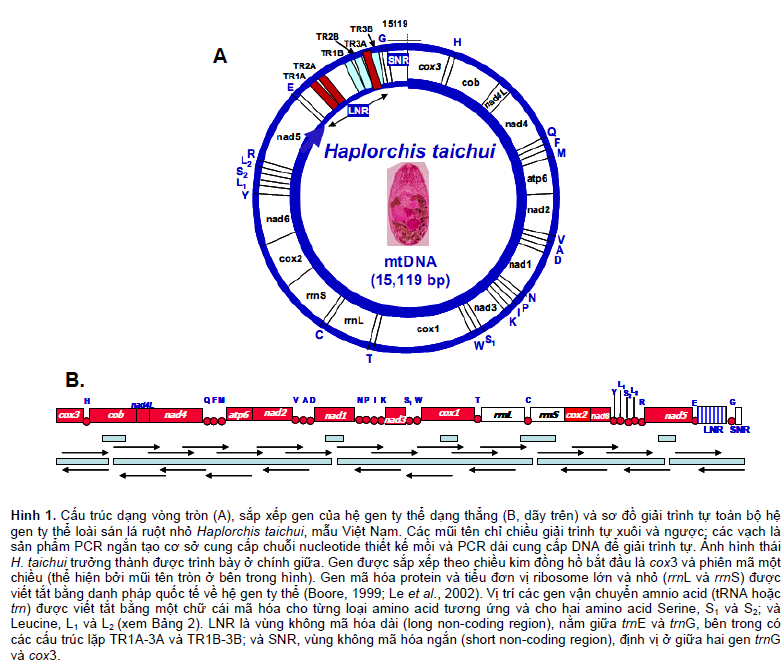
Toàn bộ chuỗi nucleotide hệ gen ty thể hoàn chỉnh (mtDNA) của sán lá ruột nhỏ H. taichui mẫu Việt Nam (HTAQT) có độ dài 15.119 bp, đã được thu nhận. Phân tích đặc điểm toàn bộ hệ gen và của từng gen cho thấy, mtDNA H. taichui có cấu trúc là vòng DNA khép kín chứa 36 gen, gồm 12 gen mã hóa protein, 2 gen RNA ribosome, 22 gen RNA vận chuyển amino acid và một vùng DNA không mã hóa. Sắp xếp thứ tự các gen được liệt kê ở Bảng 2, sắp xếp dạng vòng tròn (circular) trình bày ở Hình 1A và dạng thẳng (linear gene arangement) ở Hình 1B. Tất cả các gen ty thể đều sắp xếp cùng chiều xuôi và gần như kế tiếp nhau, chỉ cách nhau rất gần với khoảng cách giao gen (là chuỗi nucleotide nối giữa gen trước với gen liền kề sau đó) là 0- – 25 nucleotide (Bảng 2). Một số gen sử dụng 1-3 nucleotide của nhau, lồng vào nhau, để kết thúc hoặc khởi đầu gen (Bảng 2). Gen atp8 không có trong cấu trúc mtDNA của H. taichui và đó cũng là đặc trưng về cấu trúc của mtDNA ở các loài sán lá (trematode) và của tất cả các loài sán dẹt (platyhelminth) (Le et al., 2002).
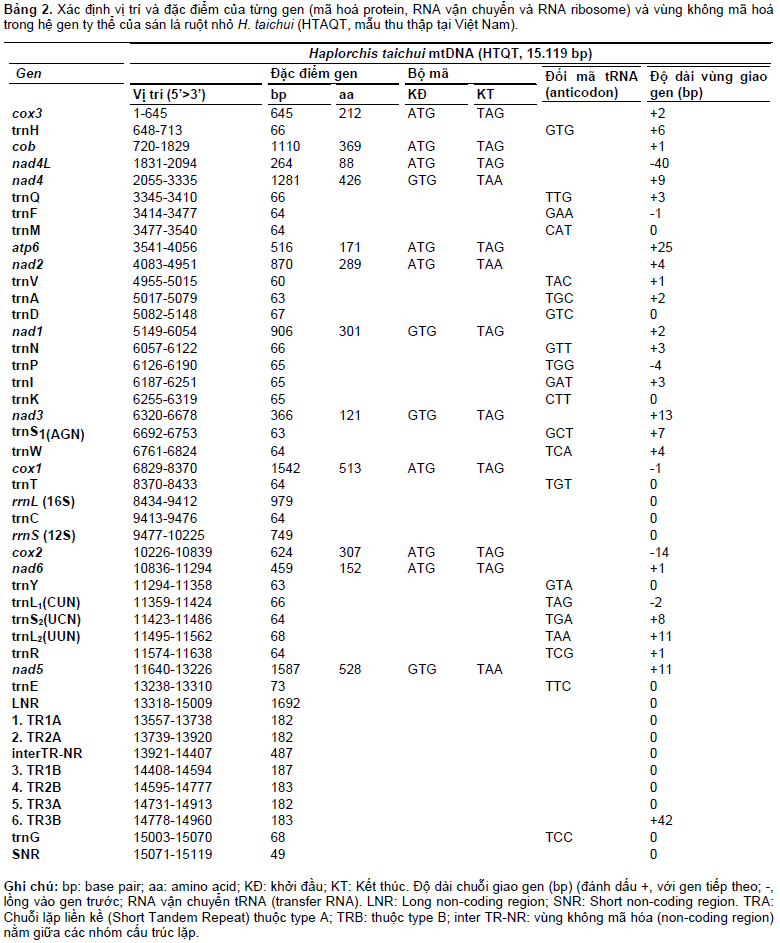
Thứ tự sắp xếp dạng thẳng (linear gene arangement) của 36 gen và vùng không mã hóa (LNR và SNR), được mở vòng tại cox3, như sau: cox3-Hcob- nad4L-nad4-Q-F-M-atp6-nad2-V-A-D-nad1-N-PI- K-nad3-S1-W-cox1-T-rrnL-C-rrnS-cox2-nad6-Y-L1- S2-L2-R-nad5-E-LNR[TR1A-3A;TR1B-3B]-G-SNR. Vùng DNA không mã hóa (NR) được chia thành 2 tiểu vùng: một tiểu vùng không mã hóa dài, LNR (long non-coding region) và một tiểu vùng không mã hóa ngắn, SNR (short). Vị trí, kích thước, đặc điểm của từng gen và vùng không mã hóa trong hệ gen ty thể của loài sán lá ruột nhỏ H. taichui HTAQT được liệt kê trong Bảng 1 và minh họa ở Hình 1. Hai gen nad4L (264 bp) và nad4 (1.278 bp) sắp xếp kế tiếp lồng vào nhau trên hai khung gen khác nhau, trong đó gen nad4 gối vào đầu 3’ của gen nad4L và chung nhau 40 nucleotide. Tương tự, gen cox2 (624 bp) và gen nad6 (459 bp) có 14 nucleotide ở đầu 3’ kết thúc của cox2 gối vào đầu 5’ khởi đầu của nad6 (Bảng 2), một trường hợp hiếm gặp ở sán dẹt được biết cho đến nay.
H. taichui sử dụng A = 19,56%, T = 39,71%, G =28,34%, C = 12,39%, để kiến tạo hệ gen ty thể (15.119 bp) với tổng thể thành phần A+T là 59,27% và G+C là 40,73%. Tổng thể thành phần A+T ở mtDNA của H. taichui cũng tương tự như ở một số loài sán lá gan lớn (fasciolid) và sán lá gan nhỏ (opisthorchiid), vào khoảng 60%, nhưng khác xa và ít hơn nhiều so với các loài sán máng họ Schistosomatidae (thường là khoảng 70% A+T) (Le et al., 2002).
Phân tích đặc điểm gen RNA ribosome và RNA vận chuyển
RNA ribosome (gồm hai tiểu phần, rrnL và rrnS) và RNA vận chuyển (gồm 22 tRNA) lần lượt, có độ dài, cấu trúc, thứ tự sắp xếp cơ bản là giống nhau giữa các loài sán dẹt, nằm xen giữa các gen mã hóa protein (Hình 1B). Hai gen RNA ribosome, rrnL và rrnS, cách nhau bằng một chuỗi gen của tRNA-Cysteine (trnC) xen vào giữa (Bảng 1). Cấu trúc bậc hai suy diễn hình “lá sồi” của tRNA ở H. taichui tương tự của các loài sán dẹt và giống như ở sán lá gan lớn F. hepatica (Le et al., 2002).
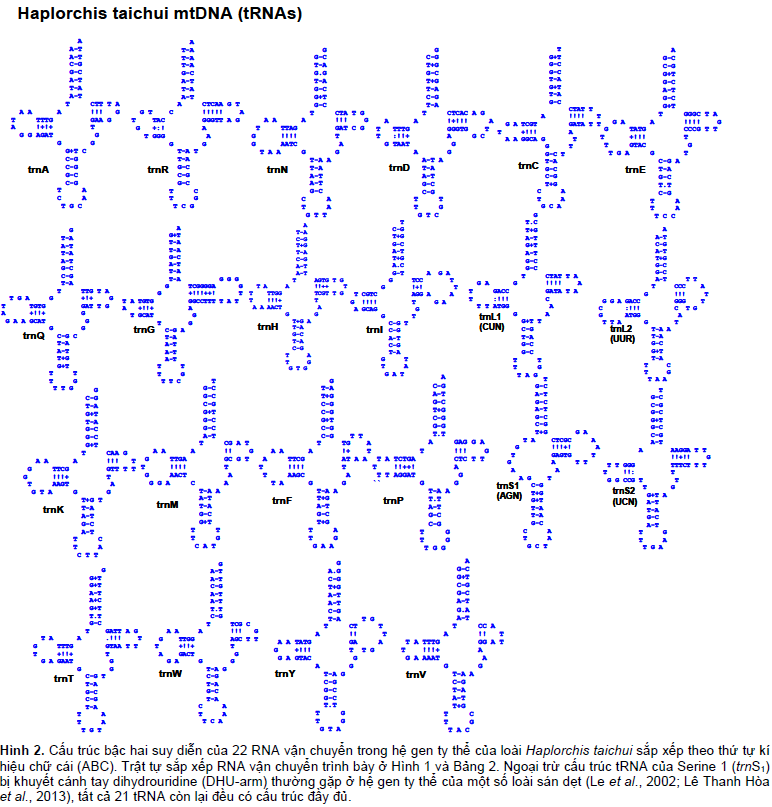
Độ dài của tRNA ở H. taichui biến động từ 60 bp (ở tRNA-Valine) đến 73 bp (ở tRNA-Glutamic acid), phần lớn tập trung vào khoảng 62- – 64 bp (Bảng 1). Vùng tiếp nhận amino acid (amino-acyl acceptor stem) chứa 7 bp (ít khi 6 bp), thường thấy ở các loài sinh vật nhân thật; vùng “biến đổi” (variable loop) nối vùng “đối mã” (anticodon) và vùng tay TΨC (TΨC-stems), chỉ chứa 4 bp (ít khi 5 hoặc 6 bp) (Hình 2). Về cấu trúc, ngoại trừ tRNA của Serine 1 (trnS1), hầu như tất cả tRNA vận chuyển (21 gen) của mtDNA H. taichui đều có cấu trúc bậc hai suy diễn hình “lá sồi” (cloverleaf) hoàn toàn đặc trưng của sán dẹt, kể cả tRNA của Serine 2 (trnS2), của Leucine (trnL1; trnL2) và của Cystein (trnC). Chỉ duy nhất cấu trúc của trnS1 của H. taichui bị khuyết cánh tay dihydrouridine (DHU-arm), tạo nên một vòm nucleotide, thường gặp ở hệ gen ty thể của một số loài sán dẹt (Le et al., 2002; Lê Thanh Hòa et al., 2013) (Hình 2).
Phân tích đặc điểm gen mã hóa protein Bảng mã di truyền được sử dụng tương tự như ở các loài sán dẹt (trematode) được nghiên cứu cho đến nay (Le et al., 2002; 2004; Lee et al., 2013). Tất cả 64 bộ mã đều được sử dụng cho mã hóa gen protein của H. taichui, trong đó bộ mã ATG (sử dụng cho 8 gen) và GTG (cho 4 gen) làm bộ mã khởi đầu; và TAA và TAG làm các bộ mã kết thúc gen mã hóa protein. Bộ mã TGA, thông thường là bộ mã kết thúc ở các loài sinh vật nhân thật (metazoan), nhưng ở sán dẹt nói chung và sán lá ruột nhỏ H. taichui trong nghiên cứu này, mã hóa cho amino acid tryptophan (W) (Le et al., 2002).
Không có bất kỳ trường hợp nào có bộ mã kết thúc khiếm khuyết (T hay TA) để kết thúc một gen như thường thấy ở hệ gen ty thể ở nhiều loài giun tròn (nematode) hoặc sinh vật nhân thật (metazoan) (Le et al., 2004). Một số bộ mã bất thường khác, như ATA hoặc ATT sử dụng làm bộ mã khởi đầu, đã có ghi nhận ở một số loài sinh vật nhân thật; và TTG hoặc GTT ở một số loài giun tròn, cũng không được tìm thấy ở loài sán lá H. taichui đang nghiên cứu ở đây (Bảng 2).
H. taichui sử dụng 10.164 bp cho mã hóa cho 3.376 amino acid kiến tạo 12 gen mã hóa protein (36 nucleotide cho 12 bộ mã kết thúc). H. taichui mtDNA sử dụng nhiều nhất, đến 301 bộ mã cho Phenylalanine (TTT-Phe), 224 cho Leucine (TTG-Leu) và 171 cho Valine (GTT-Val); và sử dụng ít nhất, chỉ có 5 bộ mã lần lượt cho Glutamine (CAA-Gln) và Arginine (CGCArg) và 6 cho Asparagine (AAC-Asn). Những bộ mã sử dụng nhiều nhất thuộc về Phenylalanine và Leucine, những bộ mã ít sử dụng nhất thường là Arginine và Glutamine (Chen et al., 2013).
Đặc điểm vùng không mã hóa protein
Vùng không mã hóa protein (non-coding region) ở mtDNA của sán lá ruột nhỏ H. taichui, được chia làm hai tiểu vùng: một tiểu vùng không mã hóa dài, LNR (long non-coding region), có độ dài 1692 bp và một tiểu vùng không mã hóa ngắn, SNR, chỉ có 49 bp, có thể coi như là một vùng giao gen (Bảng 2). Vùng LNR nằm giữa gen tRNA-Glutamic acid (trnE) và tRNAGlycine (trnG), chứa các cấu trúc “vòm” (loop) và “kẹp tóc” (hairpin) và đặc biệt, có hai nhóm cấu trúc lặp liền kề (tandem repeat, TR), chứa 3 cấu trúc giống nhau ở mỗi nhóm (Hình 1). Nhóm cấu trúc liền kề thứ nhất kí hiệu là nhóm A (type A, TR1A-TR2A-TR3A) có trình tự 182 nucleotide, đồng nhất rất cao giữa các chuỗi. Nhóm cấu trúc liền kề thứ hai kí hiệu là nhóm B (type B, TR1B-TR2B-TR3B) có trình tự 183- – 187 nucleotide, mức độ đồng nhất thấp hơn, trong đó TR3B và TR3A cùng chung nhau đến 136 bp ở mỗi chuỗi. Vùng không mã hóa có cấu trúc lặp liền kề là nét đặc trưng ở mtDNA của một số loài sán lá, như bắt gặp trong mtDNA của sán lá gan lớn F. hepatica và sán máng Schistosoma spp, tạo nên cấu trúc đa hình mang tính di truyền quần thể (Le et al., 2002).
So sánh đặc điểm hệ gen ty thể giữa các chủng H. taichui
Hệ gen ty thể của H. taichui thu thập tại Việt Nam trong nghiên cứu này được so sánh với trình tự và cấu trúc của toàn bộ hệ gen ty thể của một mẫu H. taichui mới công bố gần đây của các tác giả Hàn Quốc, được đăng kí Ngân hàng gen số KF214770 (mẫu thu từ Lào) (Lee et al., 2013). Độ dài của mtDNA H. taichui (mẫu Lào) là 15.130 bp, có trật tự sắp xếp gen và số lượng gen giống với hệ gen ty thể của H. taichui mẫu Việt Nam. Các gen mã hóa protein, gen rRNA ribosome và tRNA có độ dài và đặc điểm giống với các gen tương tự của mẫu Việt Nam. Tuy nhiên, về cấu trúc lặp liền kề, ở mẫu Việt Nam có đến 6 TR (TR1A-TR2ATR3A; và TR1B-TR2B-TR3B) đại diện đa hình (polymorphism) của hệ gen ty thể sán lá, đã được phân tích đầy đủ, nhưng ở mẫu của Lào (Lee et al., 2013) không thấy được nhắc đến. Về cấu trúc tRNA, ở mẫu H. taichui của Việt Nam chỉ có cấu trúc của trnS1, trong khi đó cả hai tRNA (trnS1 và trnS2) không có hình “lá sồi” đặc trưng mà hoàn toàn bị khuyết cánh tay dihydrouridine (DHU-arm) ở mẫu Lào trong nghiên cứu của Lee et al (2013).
Do tầm quan trọng của hệ gen ty thể trong giám định, chẩn đoán, phân loại, dịch tễ học phân tử và nghiên cứu di truyền quần thể, nên càng ngày càng có nhiều hệ gen ty thể của các loài từ bậc thấp đến bậc cao được giải mã và phân tích (O’Brien et al., 2009).
Sán dẹt trong đó có loài sán lá ruột nhỏ H. taichui, được quan tâm hàng đầu do chúng kí sinh và gây bệnh ở động vật và người, kể cả Clonorchis sinensis và Opisthorchis viverrini gây ung thư túi mật tiên phát ở người (Sripa et al., 2012). Hệ gen ty thể trình bày trong bài báo này của H. taichui có trật tự gen (gene order), cấu trúc gen (gene structure) gồm 12 gen mã hóa protein, 2 gen rRNA và 22 gen tRNA và những vùng gen không mã hóa. Ngoài những đặc điểm chung giống như một số loài sán dẹt trước đây đã được giải mã và phân tích, loài H. taichui (mẫu Việt Nam) có một số nét khác biệt đặc trưng. Ngoài bộ mã ATG, GTG cũng được sử dụng để khởi đầu và ngoài TAG, TAA dùng để kết thúc gen mã hóa protein, là đặc điểm chung về sử dụng bộ mã ở mtDNA của các loài sán dẹt được biết cho đến nay. Vùng không mã hóa phân chia thành 2 tiểu vùng, bao gồm vùng ngắn (SNR) và vùng dài (LNR), chứa 6 chuỗi lặp liền kề (TR1A-3A; và TR1B- 3B). Ngoại trừ một vài loài (sán lá gan nhỏ, O. viverrini, O. felineus, C. sinensis) không có cấu trúc lặp liền kề (Shekhovtsov et al., 2010; Cai et al., 2012), còn trong hệ gen ty thể của tất cả hơn 20 loài sán dẹt (trematode) khác đều tìm thấy chuỗi lặp TR đặc biệt này, có độ dài và số lượng thay đổi tùy loài. Tại sao có cấu trúc lặp liền kề ở vùng không mã hóa, vai trò của chúng trong tiến hóa, trong di truyền, trong chức năng của loài, vẫn là câu hỏi cần giải đáp (Bernt et al., 2013; Hammani et al., 2014). RNA vận chuyển (tRNA) trong mtDNA của loài sán lá ruột nhỏ H. taichui mới thu nhận này cũng như của các loài sán dẹt trước đây, có kích thước ngắn hơn tRNA của hệ gen nhân, có cấu trúc hình “lá sồi” đặc trưng. Bên cạnh đó, chỉ có tRNA-Serine (trnS1) có cấu trúc khuyết cánh tay DHU (DHU-arm), số lượng ít hơn so với số lượng tRNA có cấu trúc không bình thường này (2 -3 tRNA) ở các loài sán dẹt khác.
KẾT LUẬN
Hệ gen ty thể của sán lá ruột nhỏ H. taichui mẫu của Việt Nam (chủng HTAQT) đã được giải mã và phân tích đầy đủ đặc điểm gen học. Toàn bộ mtDNA của HTAQT có kích thước 15.119 bp, chứa 36 gen trong đó có 12 gen mã hóa protein (cox1, cox2, cox3, nad1, nad2, nad3, nad4L, nad4, nad5, nad6, atp6 và cob); 2 gen RNA ribosome, rrnL (16S) và rrnS (12S); 22 gen vận chuyển RNA (tRNA hay trn); và một vùng không mã hóa có các cấu trúc lặp liền kề (6 cấu trúc lặp). Đặc điểm của từng gen về độ dài, sử dụng bộ mã và phân bố, cũng như cấu trúc bậc hai của rRNA và tRNA cũng đã được xác định và phân tích đầy đủ và so sánh với mẫu của Lào do nhóm nghiên cứu Hàn Quốc phân tích.
Lê Thanh Hòa¹, Nguyễn Thị Khuê¹, Nguyễn Thị Bích Nga¹,
Đỗ Thị Roan¹, Đỗ Trung Dũng², Lê Thị Kim Xuyến¹, Đoàn Thị Thanh Hương¹
1 Viện Công nghệ sinh học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam
2 Viện Sốt rét – Ký sinh trùng – Côn trùng Trung ương, Bộ Y tế
- Mô hình nuôi vịt Grimaud trên sàn nhựa: Hướng đi mới giúp nông dân giảm chi phí và tăng năng suất chăn nuôi
- Hiệu quả của kháng sinh Tylvalosin trên heo thịt khi đối diện thử thách với chủng virus PRRS tự nhiên
- Nhà máy TĂCN thông minh: Tích hợp tự động hóa, IoT và trí tuệ nhân tạo
- Ứng dụng công nghệ chăn nuôi bò Hanwoo trong chăn nuôi bò H’Mông: Những kết quả bước đầu từ khảo sát thực địa tại Tuyên Quang
- Chủ động xây dựng hệ thống phòng bệnh lở mồm long móng: Hướng phát triển bền vững cho ngành chăn nuôi
- Bài học từ 30 năm chống cúm gia cầm H5N1 tại châu Á: Cảnh báo cho nguy cơ dịch bệnh mới nổi
- Gắn thiết bị GPS cho đàn bò: Hướng đi mới trong phát triển chăn nuôi thông minh ở vùng cao
- Đồng Nai: Tiêu hủy gần 2.000 con lợn mắc dịch tả lợn châu Phi, khẩn trương ngăn dịch lây lan
- Kiểm soát PRRS, ASF và FMD SAT1 bằng giải pháp sinh học và công nghệ chăn nuôi hiện đại – Bài học thực tiễn từ Trung Quốc
- Tổng giám đốc AVAC Việt Nam kiến nghị về diễn giải kết quả xét nghiệm ổ dịch trong bối cảnh vắc-xin ASF đã được lưu hành tự do
Tin mới nhất
T5,09/07/2026
- Mô hình nuôi vịt Grimaud trên sàn nhựa: Hướng đi mới giúp nông dân giảm chi phí và tăng năng suất chăn nuôi
- Hội Chăn nuôi Việt Nam và Trường Đại học Nông Lâm, Đại học Huế: Thúc đẩy phối hợp, chuẩn bị cho AAAP 2026
- Hướng dẫn mới về hồ sơ đăng ký xuất khẩu thực phẩm có nguồn gốc động vật sang Trung Quốc theo Nghị định 280
- Gỡ vướng thủ tục trong lĩnh vực nông nghiệp: Dabaco kiến nghị hoàn thiện chính sách về thức ăn chăn nuôi và an toàn thực phẩm
- Hiệu quả của kháng sinh Tylvalosin trên heo thịt khi đối diện thử thách với chủng virus PRRS tự nhiên
- Nhà máy TĂCN thông minh: Tích hợp tự động hóa, IoT và trí tuệ nhân tạo
- FUJINUCO kỷ niệm chặng đường 15 năm phát triển, tiếp tục mở rộng hệ sinh thái ngành chăn nuôi
- Ứng dụng công nghệ chăn nuôi bò Hanwoo trong chăn nuôi bò H’Mông: Những kết quả bước đầu từ khảo sát thực địa tại Tuyên Quang
- Hội Chăn nuôi Thỏ Việt Nam kiến nghị giảm phí kiểm soát giết mổ, gỡ khó cho ngành chăn nuôi còn nhiều dư địa phát triển
- Chăn nuôi heo thông minh: Tối ưu hóa năng suất sinh sản




-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Đón đọc Tạp chí Chăn nuôi Việt Nam số tháng 4 năm 2026
-
các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
các tạp chí đã xuất bản
-
các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
CÁC ẤN PHẨM ĐÃ XUẤT BẢN
-
Các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản
-
Các ấn phẩm đã xuất bản
-
các ấn phẩm đã xuất bản











































- Ngành chăn nuôi heo trước thách thức mới trên hành trình phát triển bền vững
- Chuyên gia bàn giải pháp sử dụng kháng sinh có kiểm soát trong chăn nuôi
- Ngành sữa Việt Nam: Cơ hội “bứt phá” từ nội lực
- Dịch tả heo châu Phi: Hiện trạng và giải pháp kiểm soát hiệu quả (Phần 1)
- Bộ NN&MT mở đợt ‘truy quét’ việc lạm dụng chất kích tăng trưởng, tăng trọng
- Cargill rút khỏi ngành thức ăn thủy sản tại Việt Nam, đóng cửa nhà máy tại Đồng Tháp và Long An
- Chăn nuôi dê bền vững theo chuỗi giá trị: Chủ nhà hàng là mắt xích quan trọng
- Da khỏe, lông đẹp: Chiến lược dinh dưỡng hiệu quả cho heo con sau cai sữa
- Lo ngại bệnh than, Campuchia ngừng nhập một số sản phẩm từ Thái Lan
- Cạn tiền, một công ty tại Nam Phi phải tiêu hủy hơn 350.000 con gà
-

[Tạp chí Chăn nuôi Việt Nam] – Theo UBND xã Minh Châu – Ba Vì – Hà Nội, xác định lợi thế về đất đai, khí hậu nên những năm qua đã đẩy mạnh sản xuất nông nghiệp là chính, trong đó xã tập trung đến phát triển chăn nuôi bò sinh sản, bò thịt. […]
-

[Tạp chí Chăn nuôi Việt Nam] – Trong bối cảnh bệnh Dịch tả lợn châu Phi (DTLCP) đã xảy ra tại TP Hồ Chí Minh, thành phố càng nỗ lực thực hiện các biện pháp kiểm soát chăn nuôi, giết mổ và tiêu thụ gắt gao. Hơn nữa, khảo sát một đêm tại thị trường […]
-

[Tạp chí Chăn nuôi Việt Nam] – Anh Trần Văn Toản, ở khu vực Bình Yên B, phường Long Hòa, quận Bình Thủy, TP. Cần Thơ là người đầu tiên ở Đồng bằng sông Cửu Long mở trang trại nuôi chim công rất thành công mà cho thu nhập hơn 200 triệu đồng/năm. 1/ […]
-

Giống chim này có khả năng thích ứng cao với điều kiện khí hậu ở nước ta, tỷ lệ nuôi sống đạt 94-99%.
-

Để đàn gà sinh trưởng phát triển tốt có tỷ lệ sống cao cần thực hiện tốt kỹ thuật úm gà con
-

Việt Nam cùng với Trung Quốc, Nhật Bản, Hàn Quốc, Mỹ… là những nước đã nuôi trồng thành công đông trùng hạ thảo.
-

Gà vảy cá được mệnh danh là “mỹ kê” đã được nhiều đại gia Việt sẵn sàng chi tiền triệu để hữu cặp gà vảy cá đẹp.
-

Cừu chính là loài vật nuôi thích hợp với những điều kiện khắc nghiệt của vùng đất Ninh Thuận.
-

Nghề nuôi chim cút đẻ hiện đang phổ biến ở rất nhiều hộ gia đình tại các địa phương và mang lại hiệu quả kinh tế khá.
-

Tỉnh Phú Thọ với địa hình đa dạng: nhiều gò, đồi thấp, dải đồng bằng thuận lợi cho chăn nuôi, trong đó, có chăn nuôi gà lông màu.












Bình luận mới nhất